摘要:为了解攀西地区蜱种类及蜱源立克次体的种类和感染状况,采用形态学和PCR方法,对随机采集自攀枝花和凉山州部分地区牛羊体表的252只蜱进行种类鉴定;利用特异性引物对立克次体ompA、gltA基因片段进行PCR扩增,随机选择阳性样本进行测序,并对测序序列进行分析,构建系统发育树。经种类鉴定,攀西地区存在微小扇头蜱和褐黄血蜱2种蜱种;蜱携带立克次体平均阳性率为29.37%(74/252),存在敬信立克次体暂定种(Candidatus Rickettsia jingxinensis)、立克次体共生菌(Rickettsia endosymbiont of Rhipilephalus microplus)、马赛立克次体(Rickettsia massiliae)共3种立克次体,其中敬信立克次体暂定种为优势种,共检出31份。不同县市的蜱携带立克次体阳性率不同,其中盐源县和昭觉县未检测到立克次体,而布拖县(57.14%)、西昌市(55.26%)阳性率较高;牛、羊体表蜱携带的立克次体阳性率分别为29.55%(26/88)和29.27%(48/164),差异不显著。结果表明,攀西地区存在多种蜱传立克次体,其分布范围较广,蜱携带率较高。结果提示,当地应做好蜱传立克次体病的防控,减少其对人和家畜的危害。立克次体是一类以蜱为主要传播媒介的病原微生物,是专性寄生于真核细胞内的革兰氏阴性原核生物,介于细菌与病毒之间,较接近于细菌,在蜱虫体内可经卵垂直传播给下一代,通过叮咬可水平传播给宿主,引起多种人兽共患病。在遗传进化史上,立克次体主要分为斑疹伤寒和斑点热群立克次体。目前我国流行的立克次体至少有10种,包括日本立克次体(Rickettsia japonica)、黑龙江立克次体(Rickettsia heilongjiangensis)、立克次体XY99(Rickettsia sp. XY99)、拉乌尔立克次体(Rickettsia raoultii)、西伯利亚立克次体BJ-90亚种(Rickettsia sibirica subsp. BJ-90)等。近几年我国又发现了几种新的蜱传立克次体,如Candidutus Rickettsia jiaonani、Rickettsia erhaii等。随着新的立克次体不断被发现,该病原逐渐受到了更多学者的重视。我国已先后从黑龙江、新疆、内蒙古、北京、福建、海南、广东、云南、广西、四川等省(区、市)的蜱中检测到立克次体。Yuan等在我国云南、广西、广东等地检测到新型立克次体GD01株,总阳性率为12.5%(71/569)。Zhang等报道我国四川省苍溪县主要流行两种立克次体,总体阳性率为33.5%(63/188)。随着调查研究深入,我国多地发现蜱携带立克次体,且其流行范围广,潜在危害大。攀西地区位于四川省西南部,山场广阔,河谷光热资源丰富,这为动植物的生长发育提供了优良环境,同时也给蜱类孳生提供了条件。本地区牛羊以放牧为主,因此增加了牛羊被蜱叮咬的概率和感染蜱传病原的风险。目前关于攀西地区蜱的区系分布及蜱传立克次体感染情况的调查研究报道较少。为了解攀西地区蜱传立克次体的流行情况,对采集自攀西地区的蜱进行种类鉴定和立克次体分子检测,以期为攀西地区蜱传立克次体病的综合防控提供依据。2021年6月至2022年3月,在凉山彝族自治州和攀枝花市,随机选择牛羊,共采集其体表的蜱252只,其中西昌市38只(羊源10只、牛源28只)、盐源县23只(牛源)、美姑县30只(羊源)、木里县19只(羊源)、会东县21只(羊源)、布拖县7只(羊源)、昭觉县7只(牛源)、普格县7只(牛源)、米易县40只(羊源)、盐边县60只(羊源),编号标记后保存于1.5 mL离心管中待检。冷冻混合球磨仪(MM400),德国莱驰公司产品;电泳仪(JY600E),北京君意电泳仪科技有限公司产品;SDS-PAGE凝胶成像系统,美国伯乐GelDocXR+公司产品;银制PCR仪(Nexus GSX)、高速离心机(5810R)、冰箱(U725)、可调微量移液枪,均为德国艾本德公司产品;试剂主要有:2×EasyTaq PCR superMix、DL 2 000 DNA Maker、TBE(tris-硼酸电泳缓冲液)、核酸染料、生理盐水等。首先对蜱进行观察,根据邓国藩等主编的《中国经济昆虫志.硬蜱科》进行初步形态学鉴定;然后将蜱虫洗净放入1.5 mL离心管中(如为成蜱可剪取一半),用干净的剪刀将蜱虫剪碎后加入200 μL灭菌生理盐水,再向离心管中加入4粒钢珠,研磨8 min及以上;取出钢珠后,按照血液/细胞/组织基因组DNA提取试剂盒说明书提取DNA并置于-80 ℃冰箱保存。根据已发表文献对蜱虫16S rRNA进行扩增,引物序列见表1。PCR扩增体系(25.0 μL):2×EasyTaq PCR superMix 12.5 μL,ddH2O 9.5 μL,引物T16SP1和T16SP2各1.0 μL,模板1.0 μL。PCR扩增程序:95 ℃ 5 min;95 ℃ 45 s、50 ℃ 45 s、72 ℃ 45 s,共38个循环;72 ℃ 6 min。使用1%琼脂糖凝胶电泳检测,所得到的阳性样本进行切胶送生物公司测序,通过序列比对确定蜱种类。
采用郭利萍报道的引物,扩增外膜蛋白A编码基因(ompA);采用高悦等报道的套式PCR引物,扩增立克次体柠檬合成酶编码基因(gltA)。以上引物均由上海杰李生物有限公司合成,引物序列见表2。
PCR扩增体系(25.0 μL):2×EasyTaq PCR superMix 12.5 μL,ddH2O 9.5 μL,上、下游引物各1.0 μL,模板1.0 μL。ompA基因扩增程序:94 ℃ 5 min;94 ℃ 30 s、46 ℃ 30 s、72 ℃ 35 s,共35个循环;72 ℃ 6 min。gltA基因第一轮扩增程序:94 ℃ 5 min;94 ℃ 30 s、50 ℃ 30 s、72 ℃ 80 s,共35个循环;72 ℃ 5 min。第二轮扩增程序:94 ℃ 5 min;94 ℃ 30 s、52 ℃ 30 s、72 ℃ 30 s,共35个循环;72 ℃ 5 min。PCR产物用1%琼脂糖凝胶电泳,选取阳性产物送至生工生物工程(成都)有限公司双向测序。将测序得到的立克次体gltA、ompA基因片段,经拼接剪切后通过Blast进行同源性比对,下载同源性较高的序列作为参考序列,同时下载不同种立克次体的同源基因序列,利用MEGA-X通过Maximum Likelihood法构建系统发育树,并设置自举值为1 000。立克次体感染情况用Excel软件分析。对不同地区、不同宿主的蜱携带立克次体阳性率,利用SPSS 21.0软件进行卡方检验。当P<0.05时,差异显著即存在统计学意义;P>0.05时,差异不显著。对252只蜱虫先进行形态观察鉴定,再通过PCR扩增16S rRNA基因,经过1%琼脂糖凝胶电泳,发现电泳条带与目的条带(460 bp)基本一致(图1)。阳性样本测序结果经Blast比对,确定为微小扇头蜱和褐黄血蜱。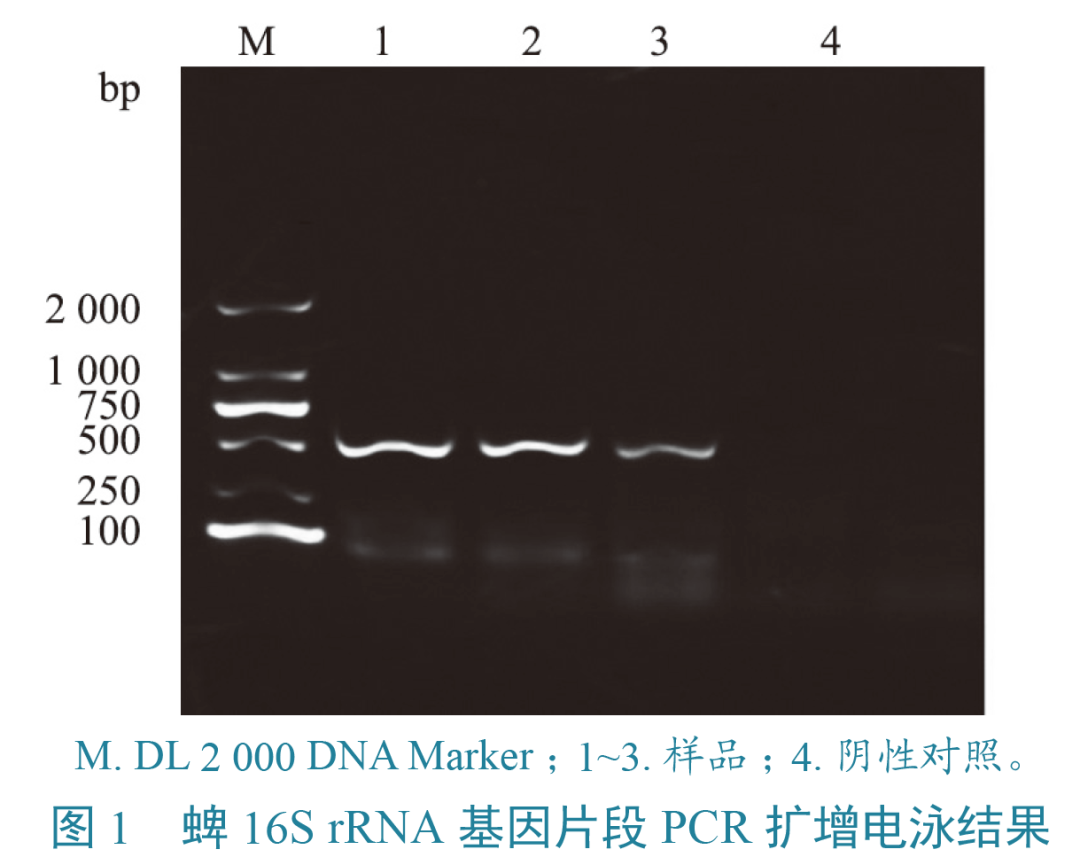
PCR扩增结果经1%琼脂糖凝胶电泳后得到的目的条带均与预期目的片段(ompA为510 bp,gltA为381 bp)大小一致(图2),分别扩增出74份ompA基因和11份gltA基因阳性样本。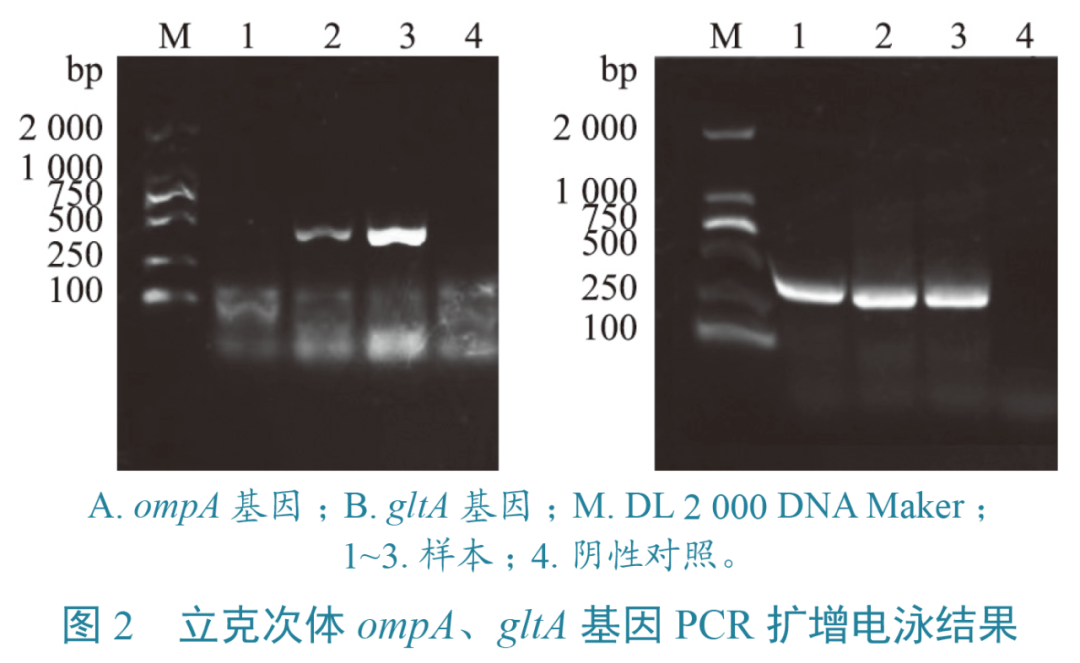
ompA基因序列分析及系统发育树构建 共有29个阳性样本ompA基因测序成功,进行剪切后序列长度约为486 bp。ompA基因片段序列同源性分析显示:主要存在两种斑点热立克次体序列,样本BTY227序列与我国陕西省长角血蜱体内检测到的敬信立克次体暂定种(Candidatus R. jingxinensis,MH932061)同源性为100%(483/483 bp),XCY98与微小扇头蜱中检测到的立克次体共生菌(Rickettsia endosymbiont of Rhipicephalus microplus,MN537556)同源性为100%(486/486 bp)。以Rickettsia akari为外群,基于ompA基因构建的系统发育树(图3)显示:BTY227与敬信立克次体暂定种聚为一支,亲缘关系近;XCY98与立克次体共生菌聚为一支,亲缘关系近,且与马赛立克次体、扇头立克次体有较近的亲缘关系。
gltA基因扩增结果分析及系统发育树构建 共有11份阳性样本gltA基因测序成功,序列长度约为366 bp。gltA基因片段序列同源性分析显示:主要存在两种斑点热立克次体序列,样本BTY227与从我国南方微小扇头蜱体内分离的敬信立克次体暂定种(Candidatus R. jingxinensis,MW114883)同源性为100%(366/366 bp),MYY86与从意大利血红扇头蜱体内分离的马赛立克次体(R. massiliae,KJ663741)同源性为99.73%(365/366 bp)。以Rickettsia bellii为外群,基于gltA基因构建的系统发育树(图4)显示:BTY227与敬信立克次体暂定种聚为一支,亲缘关系近;MYY86与马赛立克次体聚为一支,有较近的亲缘关系。
不同种类 从采集的252份蜱样品中,共检出立克次体74份,总体阳性率为29.37%(74/252)。共检出31份敬信立克次体暂定种(Candidatus R. jingxinensis),其中西昌牛源9份、米易羊源5份、普格牛源2份、会东牛源6份、美姑牛源5份、木里羊源1份、布拖羊源3份;检出立克次体共生菌(Rickettsia endosymbiont of Rhipicephalus microplus)2份,均为西昌羊源;检出马赛立克次体(R. massiliae)1份,为米易羊源。由此推断,敬信立克次体暂定种为攀西地区立克次体优势种。不同采集地 本次采集的蜱来自10个县市,不同采集地的蜱携带立克次体阳性率不同。ompA基因检测结果(表3)显示:布拖阳性率最高,为57.14%,其次为西昌55.26%,会东42.86%;经卡方检验,10个县市中,排除未检测到立克次体的盐源、昭觉后,剩余8个县市的蜱携带立克次体阳性率 差异显著(P = 0.023,<0.05)。gltA基因检测结果(表3)显示:同样是布拖阳性率最高,为28.57%,其次为普格14.29%、美姑13.33%;经卡方检验,10个县市中,排除未检出的县市,不同县市的蜱携带立克次体阳性率差异不显著(P = 0.611)。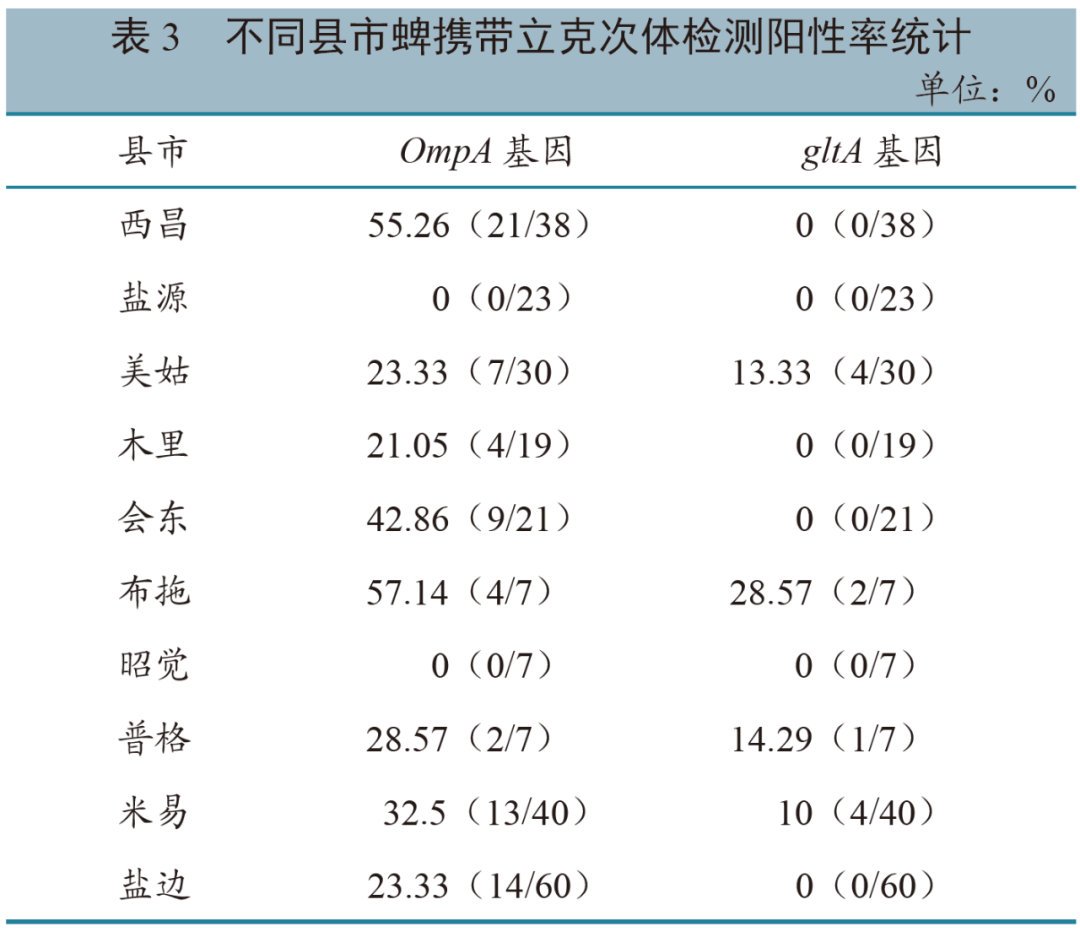
不同宿主 从总体来看,不同牛羊来源蜱携带的立克次体阳性率相近,其中牛源蜱携带的立克次体阳性率为29.55%,羊源蜱携带立克次体阳性率为29.27%(表4)。经卡方检验,牛、羊宿主蜱携带的立克次体阳性率差异不显著(P = 1.000)。但在西昌市,蜱携带的立克次体情况与宿主有显著差异(P = 0.009),而在美姑县,蜱携带立克次体情况与宿主无显著差异(P = 0.708)。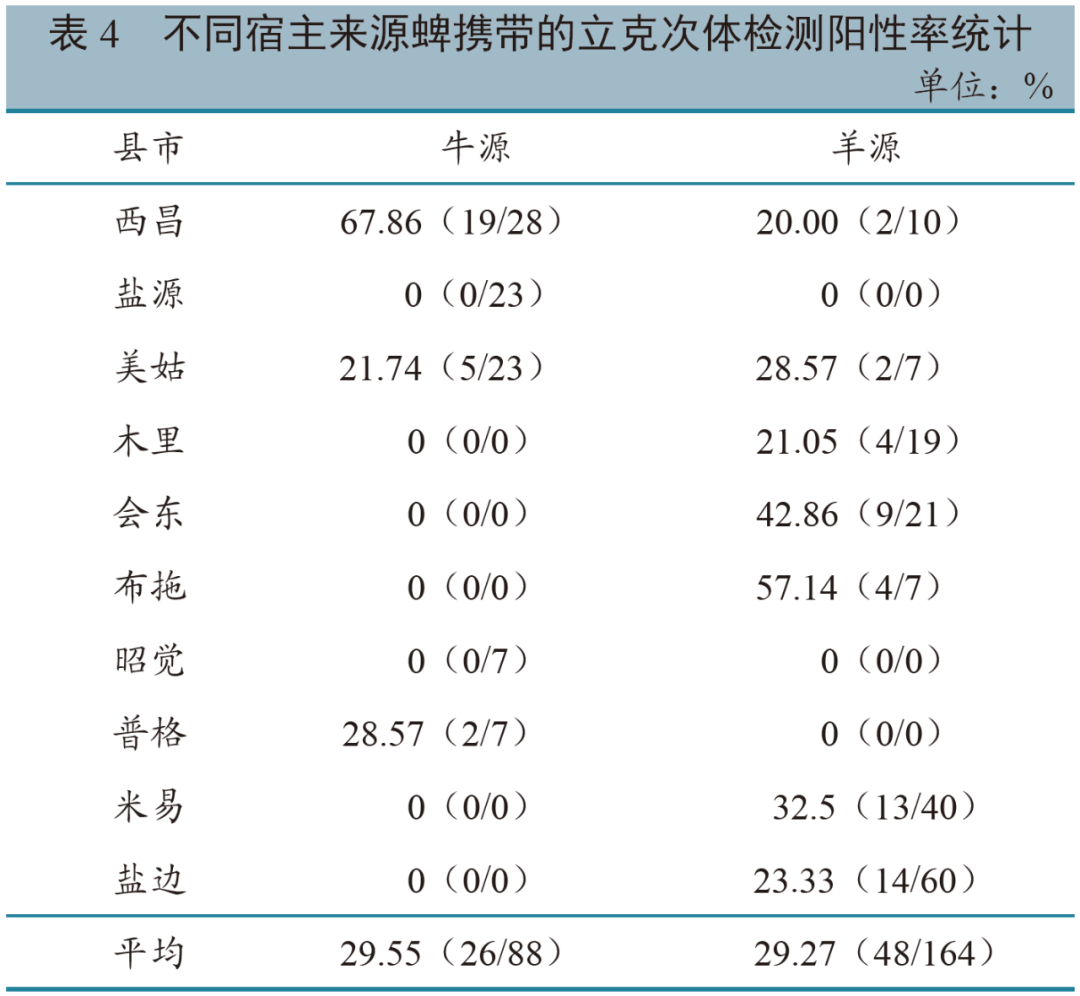
因蜱的发育经历了卵、幼蜱、若蜱、成蜱,因此通过形态学观察需要鉴定人员具有丰富的经验,因此分子学鉴定更受广大学者青睐。用于蜱分子生物学鉴定的基因主要有16S rRNA、COI、COII、ITS-1、ITS-2。16S rRNA基因序列高度保守,因此常作为蜱种类鉴定靶基因。攀西地区幅员辽阔,地理、气候等特点适合蜱生活繁衍。而该地区蜱的区系分布调查仍处于空白。本检测发现,微小扇头蜱为攀西地区的优势蜱种,并分布有褐黄血蜱。该结果与姚晓燕等对我国微小扇头蜱分布情况的总结相符。攀西地区位于川南,与云南省接壤且与其气候特点类似,也属于微小扇头蜱生长的中高适生区。目前针对蜱携带立克次体的检测方法多样,但用于流行病学调查的仍以分子生物学检测方法为主,包括普通PCR、巢式PCR、实时荧光定量PCR及自杀式PCR(suicide PCR)。主要的靶基因有16S rRNA、柠檬合成酶基因(gltA)、外膜蛋白A基因(ompA)、外膜蛋白B基因(ompB)以及细胞表面抗原1(sca1)、细胞表面抗原4(sca4)、17 kDa蛋白抗原等编码基因。其中ompA与ompB对斑点热群立克次体有较好的特异性及鉴别性,即不同斑点热群立克次体,其ompA表现出种的特异性,对斑点热群立克次体的基因型、亚型鉴定及分类具有重要意义,此外ompA基因还可以准确把握斑点热群立克次体的遗传变异特征。唐天才报道四川省石渠县斑点热立克次体总阳性率为49.48%,盛悦报道内蒙古斑点热群立克次体总阳性率为37.7%。而本研究发现,攀西地区蜱携带立克次体阳性率为29.37%,低于上述两个地区。攀西地区作为四川省的西部城市群,有发展农牧业得天独厚的优势,属于典型的半农牧地区,相比纯牧区的内蒙古及石渠县,其立克次体阳性率的差异可能与蜱种类分布及气候、海拔等自然条件不同有关。蜱虫传播需要借助宿主来完成,例如依靠鸟类迁徙、牲畜贸易等。攀西地区各地交通不便,蜱虫传播途径受限,导致各地区蜱种类及数量有差异,进而导致其携带的立克次体阳性率不同。此外,采样过程中经咨询牧民与养殖户得知,不同地区不同养殖户的驱虫意识参差不齐,导致个别地区蜱虫孳生,蜱携带的立克次体阳性率较高。不同宿主蜱携带的立克次体阳性率差异不显著,这与该地区蜱虫无差别叮咬有关,而个别地区阳性率差异显著可能与牛羊养殖模式不同等因素有关。本研究证实,攀西地区存在3种立克次体,其中敬信立克次体暂定种为优势种,其与Guo等报道的陕西省长角血蜱体内敬信立克次体暂定种序列(MH932061)同源性高达100%;且系统发育树显示,两个地区的立克次体处于同一分支。以上表明,敬信立克次体暂定种可能是国内多个地区的立克次体优势种,需要重点加强对其流行的控制。马赛立克次体与斑点热感染有关,首次于法国扇头蜱属体内被发现。Wei等首次在国内西北部的新疆伊犁县图兰扇头蜱体内检测到马赛立克次体,其蜱携带率为3.50%(4/114);Guo等在西北部塔克拉玛干沙漠图兰扇头蜱体内检测到马赛立克次体,蜱携带率为1.71%(2/117)。本检测发现,微小扇头蜱的立克次体携带率为0.40%(1/252),与上述结果基本相符,存在低携带率现象且都发现于扇头蜱属。综上,攀西地区存在发生立克次体病的潜在威胁,应重视蜱传立克次体病预防。目前攀西地区乃至凉山彝族自治州均缺乏系统的蜱种检测及蜱传立克次体的分析与研究。凉山州及周边多为少数民族群居,天然放牧为主的饲养方式使得人们与自然及牲畜接触增多,被蜱叮咬的概率较大,蜱传立克次体病发病风险较高。因此,当地应做好对牛、羊等家畜的驱虫工作,避免在蜱虫活动高峰期放牧,采取相应防护措施,防止被蜱虫叮咬。各县市应设立哨点医院,重点监测、控制和治疗蜱传立克次体病,做好宣传,提高群众防范意识,以保证公共卫生安全及畜牧业有序发展。 

